Зеленый пигмент растения. Хлорофилл — зеленый пигмент растений. Пигменты фотосинтезирующих систем Хлорофилл является важным пигментом обусловливающим зеленый цвет
Это хлорофилл. С его помощью растительность обретает соответствующий цвет. Еще в школе учат детей, что это вещество занимает важную роль в процессе фотосинтеза. Таким образом, растения не смогут существовать без него.
Но в последнее время считается, что этот пигмент можно использовать для здоровья человека. Существует информация, что продается в аптеке, приобретение его не составляет большого труда. Считается, что он сможет помочь в лечении многих болезней. Но обладает ли на самом деле это вещество целительными свойствами?
Уже говорилось, что хлорофилл - зеленый пигмент растения, придающий ему соответствующую окраску. Это важный элемент в жизнедеятельности растительности, требующийся для фотосинтеза. Хлорофилл имеет особый химический состав: атом магния окружают атомы азота, водорода, углерода и кислорода.
Почти сотню лет назад Ханс Фишер сделал удивительное открытие. Он заметил, что имеют сходство химические структуры хлорофилла и гемоглобина. Отличие состоит в том, что вместо магния гемоглобин содержит железо. Из-за этого пигмент хлорофилл начали называть кровью растений. Многих ученых заинтересовало это вещество, они начали его исследовать. Некоторым захотелось использовать его в медицине.
Использование хлорофилла
Зеленый пигмент растения на сегодня применяют в качестве пищевой добавки. Она больше известна как Е-140. С ее помощью заменяют красители, которые используют для Производным хлорофилла является тринатриевая соль. Ее используют в промышленности пищевых продуктов в качестве красителя, названа она Е-141.
Не могли ученые на то, что структура гемоглобина так похожа на хлорофилл. Из-за этого его применяют не только для пищевых добавок. На сегодняшний день выпускается экстракт зеленого пигмента. Его называют жидкий хлорофилл и применяют в медицине в качестве целительного средства. Но полезен ли он на самом деле?
Обещания производителей относительно жидкого хлорофилла
На сегодня привлекает интерес к себе жидкий хлорофилл. Растение содержит зеленый пигмент, который используется для этой биологической добавки. Средство привлекло людей, желающих поправить здоровье. Производитель, который его выпускает, считает, что препарат благотворно влияет на организм, так как строение пигмента очень схоже с гемоглобином.

Покупателям рассказывается информация, что жидкий хлорофилл имеет такие свойства:
- Выводит шлаки, токсины из организма.
- Регулирует уровень гормонов, которые находятся в крови.
- С ним кислотно-щелочной баланс всегда будет в норме.
- Кровь насыщается минералами, полезными веществами, витаминами.
- Регенерация тканей, происходят быстрее.
- Иммунитет улучшается.
- Он способен помочь в некоторых гинекологических патологиях.
Мнение специалистов
Эта пищевая добавка представлена как происхождения, который способен оказать чрезвычайное лечебное воздействие. С его помощью можно лечить болезни, а также заниматься профилактикой. Но что думают об этом специалисты?
Мнения врачей разделились:
- Противники предполагают, что применять жидкий хлорофилл - это бессмысленное дело из-за того, что вещество не способно усвоиться в организме человека полноценно. Теории про целительные свойства они также опровергают.
- Но есть специалисты, которые подтверждают некоторые лечебные свойства препарата. Они заметили, что он действительно выводит шлаки, укрепляет иммунную и сердечно-сосудистую системы.
Однозначного мнения не существует. Из-за этого каждый человек самостоятельно решает, нужно ли ему это средство. Но, кроме этого, зеленый пигмент растения нужен для очищения воздуха, что важно для жизни человека.
Фотосинтез
Одно точно известно, что помочь насытить воздух кислородом может хлорофилл. Фотосинтез - сложный процесс, где участвуют растения, энергия солнца. Происходит химическая реакция, с помощью которой из углекислого газа появляется кислород. Только этот процесс жизнедеятельности всего на планете использует энергию солнца.

Улавливают солнечный свет фотоавтотрофы. Такой процесс происходит в растениях, в некоторых водорослях и одноклеточных. Несмотря на то что фотосинтез осуществляют низшие жизненные сущности, половина работы выпадает на растения.
Наземные представители растительности получают воду через корни, которая необходима для данного процесса. На поверхности листьев есть небольшие отверстия, через которых поступает углекислота. В процессе всего этого освобождается кислород. Без хлорофилла этот процесс невозможен, так как именно этот зеленый пигмент растения поглощает солнечную энергию.
Хотя существует и бесхлорофилльный фотосинтез. Он был замечен у солелюбивых бактерий, вмещающих светочувствительный фиолетовый пигмент. Последний способен поглощать свет. Но это единичный случай. В основном участие принимает хлорофилл.
Свойства хлорофилла, открытые наукой
Зеленый пигмент начали плотно изучать в науке. Было доказано, что жидкий хлорофилл способствует регенерации клеток. Но сделать мощный антибиотик все-таки не удалось, поэтому отдавалось предпочтение таблеткам.
Но большого прогресса достигли исследования в стоматологии. Заинтересовавшись целебными свойствами хлорофилла, его изучали, заметили положительное влияние на ротовую полость. Роберт Нар изобрел программу, с помощью которой можно было бороться с кариесом. Была выпущена зубная паста, в составе которой был хлорофилл. Как известно, этот зеленый пигмент активно участвует в фотосинтезе, с помощью которого производится кислород. А это мощный агент, устраняющий бактерии, в том числе и те, которые провоцируют кариес. Из-за этого паста заслужила признание, так как показала отличный результат.
Также были положительными исследования, с помощью которых выявилось, что пигмент борется с панкреатитом, если принимать его внутрь.

Итак, хлорофилл играет важную роль в жизни не только растений, а и всех людей. С его помощью происходит фотосинтез, выделяется кислород, необходимый человеку. Также жидкий хлорофилл начали применять в медицине. Многие исследования показали высокий результат.
Хлорофилл А против В
Растения и водоросли - это живые организмы, которые могут создавать свою собственную пищу, а животные получают свою пищу от этих растений. Этот процесс создания пищи называется фотосинтезом и использует хлорофилл. Хлорофилл является зеленым пигментом в растениях и водорослях, который по существу используется при фотосинтезе. Он поглощает свет и энергию от синей и красной частей электромагнитного спектра, но не поглощает лунку зеленой порции, которая придает растениям хлорофилл в растениях свой зеленый цвет Затем свет и энергия переносятся в реакционные центры двух фотосистем, Photosystem I и Photosystem II. Эти фотосистемы имеют реакционные центры, P680 и P700, которые поглощают и используют энергию, которую они получают от других пигментов хлорофилла. Фотосинтез использует два типа хлорофилла, хлорофилла а и б, для получения энергии. Хлорофилл А Хлорофилл а поглощает энергию от длин волн сине-фиолетового и оранжево-красного света при 675 нм. Он отражает зеленый свет, который придает хлорофиллу свой зеленый вид. Это очень важно в энергетической фазе фотосинтеза, потому что молекулы хлорофилла a необходимы до того, как фотосинтез может продолжаться. Это первичный фотосинтетический пигмент. Это реакционный центр антенной решетки, который состоит из основных белков, которые связывают хлорофилл а с каротиноидами. Организмы, в частности кислородные фотосинтезирующие, используют хлорофилл а и используют различные ферменты для биосинтеза. Хлорофилл B Хлорофилл б поглощает энергию от длин волн зеленого света при 640 нм. Это вспомогательный пигмент, который собирает энергию и передает его на хлорофилл а. Он также регулирует размер антенны и более абсорбируется, чем хлорофилл a. Хлорофилл b дополняет хлорофилл a. Его добавление к хлорофиллу а увеличивает спектр поглощения за счет увеличения диапазона длин волн и расширения спектра поглощаемого света. Когда имеется мало света, растения производят больше хлорофилла b, чем хлорофилл a, чтобы увеличить его способность к фотосинтезу. Это необходимо, потому что молекулы хлорофилла a захватывают ограниченную длину волны, поэтому вспомогательные пигменты, такие как хлорофилл b, необходимы для захвата более широкого диапазона света. Затем он передает захваченный свет от одного пигмента к другому, пока не достигнет хлорофилла а в реакционном центре. Хлорофилл а не может эффективно функционировать без помощи хлорофилла b, а хлорофилл b не может эффективно производить достаточно энергии самостоятельно. Эти два типа хлорофиллов поэтому очень важны в процессе фотосинтеза. Они работают лучше всего вместе. Резюме 1. Хлорофилл а является основным фотосинтетическим пигментом, а хлорофилл б является вспомогательным пигментом, который накапливает энергию и передает его хлорофиллу а. 2. Хлорофилл а поглощает энергию от длин волн сине-фиолетового и оранжево-красного света, а хлорофилл б поглощает энергию от длин волн зеленого света. 3. Хлорофилл а поглощает энергию при 675 нм, тогда как хлорофилл б поглощает энергию при 640 нм. 4. Хлорофилл b является более абсорбирующим, а хлорофилл a - нет. 5. Хлорофилл а является реакционным центром антенной решетки основных белков, а хлорофилл б регулирует размер антенны.
Уже давно было подмечено, что лишенные окраски клетки и органы растения (например, корни) не способны фотосинтезировать. Особенно наглядны в этом отношении альбиносы (от лат. альбус - белый) - неокрашенные проростки, развивающиеся иногда у высших растений даже при хорошем освещении. Отсутствие окраски вызывается у таких растений какими-то внутренними факторами, препятствующими образованию особых красящих веществ - пигментов (от лат. пигментум - краска) при самых благоприятных для этого внешних условиях. При полном альбинизме, который довольно часто можно наблюдать у кукурузы, проростки не способны осуществлять фотосинтез и поэтому погибают от истощения вскоре после появления двух или трех листочков. Частичный альбинизм обычно встречается при пестролистности, когда белыми и не способными к фотосинтезу оказываются отдельные участки листа. Пестролистные формы отличаются своеобразной красотой и в связи с этим широко используются как декоративные растения.
Явление альбинизма у растений свидетельствует об огромной роли пигментов для фотосинтетической деятельности. Установлено, что процесс фотосинтеза связан прежде всего с поглощением света пигментами пластид. Энергия солнечной радиации сможет превратиться в химическую энергию образующихся при этом органических веществ только в том случае, если она сначала будет поглощена пигментами фотосинтезирующих систем и в первую очередь зеленым пигментом - хлорофиллом.
Пигменты фотосинтезирующих систем можно разделить на три основные группы: хлорофиллы, каротиноиды и фикобилипротеиды. Пигментный состав различных фотосинтезирующих систем неодинаков и зависит от принадлежности растения к той или иной систематической группе.
Важнейшие представители пигментов фотосинтезирующих систем у различных групп растений
| Пигменты | Серобактерии | Водоросли | Высшие растения | |||||
| пурпурные | зеленые | сине-зеленые | красные | диатомовые | бурые | зеленые | ||
| Хлорофиллы | ||||||||
| хлорофилл a | - | - | + | + | + | + | + | + |
| хлорофилл b | - | - | - | - | - | - | + | + |
| хлорофилл c | - | - | - | - | + | + | - | - |
| хлорофилл d | - | - | - | + | - | - | - | - |
| бактериохлорофилл | + | - | - | - | - | - | - | - |
| бактериовиридин | - | + | - | - | - | - | - | - |
| Фикобилипротеиды | ||||||||
| фикоэритрин | - | - | + | + | - | - | - | - |
| фикоциан | - | + | + | - | - | - | - | |
| Каротиноиды | ||||||||
| β-каротин | - | - | + | + | + | + | + | + |
| лютеин | + | - | - | + | + | |||
| виолаксантии | - | - | - | + | + | + | ||
| фукоксантин | - | - | - | ? | + | + | - | - |
Условные обозначения:
+ наличие пигмента; - отсутствие; ? - очень малое количество; пустые клетки - не было исследований.
Для обозначения различных форм хлорофилла используются буквы латинского алфавита: а, b , с, d .
Из таблицы видно, что среди различных форм хлорофилла наибольшее распространение имеет хлорофилл a - обязательный компонент пластид всех фотосинтезирующих растений. Хлорофилл a отсутствует у пурпурных и зеленых серобактерий, для которых характерна наиболее примитивная форма синтеза органических веществ, так называемый бактериальный фотосинтез или фоторедукция. Эти бактерии восстанавливают CO 2 в анаэробных условиях до уровня органических веществ за счет энергии солнечного света с использованием сероводорода или некоторых других неорганических веществ, при этом не происходит выделения молекулярного кислорода. Вместо хлорофилла a пурпурные серобактерии содержат бактериохлорофилл, а зеленые серобактерии - бактериовиридин.
Строение пигментов
Изучение химических свойств хлорофилла a позволило расшифровать его структуру. Правильность предложенной структурной формулы подтверждена полным синтезом хлорофилла, осуществленным в 1960 г. почти одновременно в лабораториях М. Штрелля и Р. Вудварда. Основу молекулы хлорофилла составляют четыре пиррольных кольца (I, II, III и IV). Пиррол представляет собой пятичленное циклическое соединение. Остатки пиррольных колец, соединяясь между собой, образуют ядро молекулы хлорофилла, в центре которого помещается атом магния. Он соединяется с азотом пиррольных колец, причем два атома азота соединяются с магнием своими основными валентностями, а два других - дополнительными валентностями. Зеленый цвет хлорофилла как раз и определяется наличием металла в его молекуле. В этом легко убедиться, стоит только осторожно подействовать на спиртовую вытяжку хлорофилла кислотой. Магний замещается двумя атомами водорода из кислоты и получается бурый продукт - феофитин. Восстановить зеленый цвет вытяжки можно путем добавления какой-либо соли магния или другого двухвалентного металла.
Одной из специфических черт хлорофиллов является наличие в их молекуле добавочного кольца из пяти углеродных атомов (V). Кислотная, или карбоксильная, группа (-СООН) этого кольца соединена эфирной связью с метиловым спиртом (СН 3 ОН). Вторая карбоксильная группа, находящаяся около четвертого пиррольного кольца, соединена эфирной связью с другим спиртом - фитолом (С 20 Н 39 ОН).
Таким образом, по своей химической природе хлорофилл (см. его структурную формулу на обложке книги) - сложный эфир двухосновной кислоты хлорофиллина, в одном карбоксиле которой водород замещен остатком метилового спирта (-СН 3), а в другом - остатком фитола (-С 20 Н 39).
Интересно отметить, что наличие остатка фитола обусловливает липофильность молекулы хлорофилла, проявляющуюся в его растворимости в жировых растворителях. Кроме того, остаток фитола играет существенную роль в образовании комплексов хлорофилла с липоидами. Выделенный в чистом виде фитол напоминает бесцветную воскообразную массу.

По своей химической природе хлорофилл весьма близок к некоторым важным дыхательным ферментам, а также к красящему веществу крови - гему гемоглобина. Общность химической природы двух основных пигментов живых организмов, а именно хлорофилла растений и гемоглобина животных, заключается в том, что у гема также имеется ядро, состоящее из четырех соединенных друг с другом пиррольных колец, в центре которого находится атом металла, только у хлорофилла - это магний, а у гема - железо. Исследования С. Граника, Т. Н. Годнева и других ученых показали, что биосинтез хлорофилла и гема осуществляется на начальных этапах аналогичным путем из одних и тех же веществ. Эти факты сходства в строении в способе образования хлорофилла и гема гемоглобина еще раз указывают на родство происхождения столь отдаленных организмов, как растения и животные.
Помимо хлорофилла a, обладающего сине-зеленой окраской, в хлоропластах зеленых водорослей и высших растений обнаружена еще другая форма желто-зеленого цвета - хлорофилл b. В отличие от хлорофилла a, имеющего эмпирическую формулу C 55 H 72 O 5 N 4 Mg, хлорофилл b содержит лишь на два атома водорода меньше и на один атом кислорода больше. В структурном отношении это незначительное различие сводится к тому, что вместо метильной группы (-СН 3) при втором пиррольном кольце у хлорофилла b имеется альдегидная группа COH. Красные водоросли содержат два зеленых пигмента: хлорофиллы a и d, очень близкие по структуре. Они различаются только тем, что у хлорофилла d при первом пиррольном кольце стоит не винильная группа (-СН = СН 2), а альдегидная. У бурых и диатомовых водорослей вместо хлорофилла b содержится хлорофилл с, который в отличие от других форм лишен остатка фитола.
Пурпурные серобактерии имеют весьма близкий к хлорофиллу a бактериохлорофилл. Небольшое различие состоит в том, что у этого пигмента при первом пиррольном кольце находится вместо винильной ацетильная группа (-СО-СН 3), а также на два атома водорода больше во втором пиррольном кольце. Характерный для зеленых серобактерий бактериовиридин (иногда его называют хлоробиум-хлорофиллом) изучен пока недостаточно полно, состав и строение окончательно еще не установлены. Известно только, что он не дает характерных реакций на присутствие в его молекуле пятого кольца и содержит вместо фитола транс-фарнезол (C 15 H 26 O).
Наряду с хлорофиллами к постоянным компонентам комплекса пигментов фотосинтезирующих систем относятся каротиноиды. Обычно это желтые, оранжевые и светло-красные пигменты, которые хорошо растворяются в жирах и растворителях жиров, но не растворяются в воде. Как правило, их окраска маскируется интенсивным зеленым цветом хлорофиллов. Поэтому в природе только осенью, когда преимущественно происходит разрушение зеленых пигментов, листья желтеют и каротиноиды становятся доступными для непосредственного наблюдения.
К основным каротиноидам пластид относятся каротины, представляющие собой ненасыщенные углеводороды (эмпирическая формула C 40 H 56), и их кислородсодержащие производные - ксантофиллы.
Молекулы большинства каротиноидов имеют вид длинной углеродной цепи, которая часто заканчивается на одном или на обоих концах кольцом ионона. Атомы углерода соединяются друг с другом в длинную цепь посредством чередующихся одинарных и двойных связей. Такие связи называются конъюгированными, их наличие в молекуле обусловливает характерную окраску каротиноидов.
Пластиды высших растений и большинства водорослей содержат до восьми и больше различных каротиноидов. Кратко остановимся на характеристике некоторых из них.
Типичными, широко распространенными каротиноидами высших растений являются β-каротин, лютеин и виолаксантин. β-каротин - это желто-оранжевый пигмент. Название его происходит от латинского слова карота - морковь, в корнях которой отлагается значительное количество его в виде желто-оранжевых кристаллов. К β-каротину близок по составу и структуре лютеин. Это желтый пигмент из группы ксантофиллов, представляющий собой двухатомный спирт с эмпирической формулой С 40 Н 56 О 2 , причем обе спиртовые группы (-ОН) находятся в кольцах ионона. Другим близким к лютеину пигментом из группы ксантофиллов является виолаксантин с эмпирической формулой С 40 Н 56 О 4 . Он имеет в своем составе на два атома кислорода больше, чем лютеин. Установлено, что эти два атома кислорода входят в состав иононовых колец в виде добавочных эпокси-группировок (> O), отличающихся большой лабильностью и реакционной способностью.
К группе ксантофиллов относится также и фукоксантин (от лат. фукус - род бурых водорослей и от греч. ксантос - желтый), встречающийся у бурых и диатомовых водорослей. Он имеет оранжево-бурый цвет, близок по своим свойствам к лютеину и виолаксантину, однако состав и структура его окончательно еще не выяснены, поэтому в эмпирической формуле (С 40 Н 54-60 О 6) указывается возможное количество атомов водорода.
Набор каротиноидов в хроматофорах фоторедукторов (пурпурные и зеленые серобактерии) существенно отличается от такового в хлоропластах фотосинтетиков. Для фоторедукторов характерно отсутствие пигментов из группы ксантофиллов, содержащих эпокси-группировки. Основным каротиноидом зеленых серобактерий оказался желтый пигмент, γ-каротин, у которого имеется только одно иононовое кольцо. Что касается пурпурных серобактерий, то у них широко распространены ликопин и другие близкие к нему каротиноиды ациклического типа, не имеющие в своем составе ни одного иононового кольца.
Помимо хлорофиллов и каротиноидов, красные и сине-зеленые водоросли содержат еще дополнительные пигменты, хорошо растворимые в воде, так называемые фикобилипротеиды, к которым относятся фикоэритрины (красного цвета) и фикоцианы (сине-фиолетового цвета). Само название указывает на то, что в химическом отношении эти пигменты представляют собой сложные белки - протеиды. Небелковыми компонентами у них служат обусловливающие окраску фикобилины, которые связываются с белками в сложные комплексы. Связь небелковой группы с белком настолько прочна, что отделить фикобилины от белка можно лишь путем кипячения с сильными кислотами.

Небелковым компонентом у фикоэритринов является мезобилиэритрин, а у фикопианов - мезобиливиолин, структура которых представлена на рисунке. Как видно, фикобилины имеют в своем составе четыре пиррольных кольца, соединенных друг с другом в виде развернутой цепи, однако в деталях структура их окончательно еще не установлена. Важно подчеркнуть, что фикобилины весьма близки по своей химической природе к желчным пигментам животных.
Теперь кратко остановимся на вопросе о количественном содержании основных представителей различных групп пигментов и о их соотношении. Общее количество хлорофилла невелико и составляет в среднем около 1 % от сухого вещества листа, но оно может сильно варьировать, изменяясь у водорослей от 0,6 до 1,5%, а у высших наземных растений от 0,7 до 1,3%. Даже в пределах одного и того же растения разные листья часто значительно отличаются по содержанию зеленых пигментов, Хрофилла а всегда больше, на три его молекулы обычно приходится одна молекула хлорофилла b. Однако у теневых листьев и теневыносливых растений хлорофилла b больше, чем у светолюбивых. Так, у многих зеленых водорослей, произрастающих в условиях сильного затенения, среднее отношение хлорофилла a к хлорофиллу b снижается до 1,4. Вместе с тем их соотношение у исключительно светолюбивых растений в среднем доходит до 5,5.
В нормально развитых листьях каротиноидов гораздо меньше, чем. зеленых пигментов, примерно в 3-5 раз, а иногда даже еще меньше. Отношение же ксантофиллов к каротину может изменяться в зависимости от вида растений и условий их произрастания в очень широких пределах, от 1,8 до 15, но в среднем оно таково, что на 2-3 молекулы ксантофиллов приходится одна молекула каротина.
По данным В. Н. Люб именно, отношение фикоэритрина к хлорофиллу колеблется у разных видов красных водорослей от 0,06 до 0,66. Содержание фикоэритрина возрастает с увеличением глубины обитания этих водорослей. В среднем на одну квантосому приходится 230 молекул хлорофилла и 48 молекул каротиноидов.
Оптические свойства и состояние пигментов
Глубоко понять механизм процесса фотосинтеза возможно лишь на основе современных представлений о физических свойствах пигментов и их состоянии в живой клетке. Основная функция пигментов фотосинтезирующих систем заключается в поглощении света. Отсюда понятно то исключительно важное значение, какое имеет познание оптических свойств различных пигментов и в первую очередь их спектров поглощения.
Поглощение световой энергии пигментами осуществляется не сплошным, а избирательным образом и зависит от структуры молекулы данного пигмента. Явление это может наблюдаться как в видимой части солнечного спектра, так и в невидимых участках (инфракрасном и ультрафиолетовом), однако особый интерес представляют спектры поглощения пигментов только в интервале фотосинтетически активной радиации (380-720 ∙ 10 -9 м).
Хлорофиллы a, b, с, d, а также бактериовиридин имеют два основных максимума поглощения в красной и сине-фиолетовой частях спектра с незначительными различиями в их положении. Как видно на рисунке, у хлорофилла b максимум поглощения в красной части спектра несколько смещен в сторону коротковолновых лучей, а в сине-фиолетовой части, наоборот, в сторону длинноволновых лучей по сравнению с положением аналогичных максимумов у хлорофилла а.
Спектры поглощения растворов зеленых пигментов показывают, что сине-фиолетовые и красные лучи поглощаются интенсивно, тогда как зеленые и желтые - очень слабо, поэтому последние хорошо проходят через эти пигменты, обусловливая их цвет. Поскольку желтые лучи маскируются зелеными, окраска хлорофилла получается зеленая. Однако тот же самый раствор хлорофилла, имеющий в проходящих лучах зеленую окраску, в отраженном свете воспринимается как вишнево-красный благодаря явлению флуоресценции.

Флуоресценция (от англ. флуэреснт - свечение) представляет собой кратковременное свечение, прекращающееся практически одновременно с прекращением освещения данного вещества. Способность к такого рода свечению является очень важным оптическим свойством зеленых пигментов, указывающим на их значительную фотохимическую активность. Это свойство гораздо лучше обнаруживается не на живых листьях, а на растворах зеленых пигментов, потому что свечение хлорофилла, находящегося в клетках листа, примерно в 10 раз слабее, чем в растворе. Красное свечение хлорофилла легко наблюдать, если рассмотреть его вытяжку (например, спиртовую) со стороны падающего света, а за пробиркой с вытяжкой поместить черный фон, поглощающий все проходящие через нее лучи. При этом естественно возникает вопрос: почему окраска вытяжки хлорофилла в отраженном свете не такая, как в проходящем? Постараемся кратко объяснить это.
Свет поглощается дискретно, т. е. порциями в виде квантов, или фотонов. Энергию кванта (Е) можно выразить, как E = h ∙ v, где h - постоянная Планка, v - частота колебаний света, обратно пропорциональная длине волны. В основном, устойчивом состоянии молекулы хлорофилла каждый электрон находится на определенном энергетическом уровне и обладает соответствующим запасом энергии. Поглощение кванта света приводит к переходу электрона на более высокий энергетический уровень, при этом молекула становится возбужденной с избыточным количеством энергии. Однако время, в течение которого молекула существует в возбужденном состоянии, чрезвычайно мало, примерно 10 -9 сек, а следовательно, возбужденное состояние неустойчиво и вскоре происходит возвращение молекулы к более устойчивому состоянию с меньшим запасом энергии, или к исходному. Возбужденная молекула может терять энергию несколькими путями, наиболее важным из которых является путь превращения этой энергии в химическую энергию. Кроме того, энергия возбужденной молекулы может либо рассеиваться в виде тепловой энергии как внутримолекулярным, так и межмолекулярным путем, либо испускаться в виде кванта света. При флуоресценции возбужденные молекулы испускают кванты света, обладающие меньшей энергией, а значит, большей длиной волны по сравнению с теми квантами, которые вызывают это свечение. Каждое вещество испускает лучи определенной длины волны, т. е. имеет свой специфический спектр флуоресценции. По таким спектрам можно довольно легко отличить одно флуоресцирующее вещество от другого.
Спектр флуоресценции хлорофилла имеет только один основной максимум свечения, приходящийся на более длинноволновую красную область спектра в сравнении с максимумом поглощения в этой же области. Это значит, что при флуоресценции хлорофилл всегда испускает только длинноволновые красные лучи даже в том случае, когда он освещается синими или фиолетовыми лучами. Вот почему вытяжка хлорофилла имеет в отраженном свете темно-красную окраску.
Кроме зеленых пигментов, флуоресцировать способны и фикобилипротеиды, причем фикоциан в растворе обнаруживает красное свечение, фикоэритрин - оранжевое. Каротиноиды же обычно считаются не способными к флуоресценции.
Ученых давно интересовало, в каком состоянии находится хлорофилл в хлоропласте. Особое состояние пигментов в живой клетке убедительно доказывается, в частности, несовпадением их оптических свойств со свойствами растворов тех же самых пигментов, например несовпадением спектров поглощения, а также спектров флуоресценции. Максимумы поглощения хлорофиллов и ряда других пигментов, находящихся в живой клетке, оказываются сильно сдвинутыми в длинноволновую область по сравнению с максимумами поглощения света растворами этих пигментов. Так, раствор бактериохлорофилла имеет в инфракрасной области спектра только один максимум поглощения при 770-780 ∙ 10 -9 м (7700-7800 Å), тогда как этот же пигмент, но находящийся в клетках пурпурных серобактерий, обнаруживает три максимума поглощения, причем все они приходятся на более длинноволновый участок инфракрасных лучей: 800, 850 и 890 ∙ 10 -9 м (8000, 8500 и 8900 Å). Максимум флуоресценции бактериохлорофилла живых клеток также значительно сдвинут в длинноволновую часть и приходится на 900-920 ∙ 10 -9 м вместо 780-790 ∙ 10 -9 м - положения максимума флуоресценции этого пигмента в растворе.
До недавнего времени широкое распространение имело положение, высказанное крупнейшим специалистом в области фотосинтеза В. Н. Любименко, что особое состояние зеленых пигментов в живой клетке обусловливается связью их с белками. Однако сейчас накопилось достаточно экспериментальных данных, свидетельствующих о том, что специфику оптических свойств хлорофиллов живых клеток недостаточно объяснить только связью их с белками. А. А. Красновский, Т. Н. Годнев и другие установили возможность существования одного и того же пигмента живой клетки в нескольких формах, причем некоторым из них свойственно агрегированное состояние с разным характером упаковки молекул. Оказалось, что положение максимумов поглощения на спектрах хлорофилла а, бактериохлорофилла и бактериовиридина зависит от характера агрегации. В качестве примера, иллюстрирующего влияние агрегации пигмента на его спектры, приведем результаты опытов по изучению бактериохлорофилла, находящегося в живых клетках (пурпурные бактерии) и модельных системах; эти опыты проведены А. А. Красновским с сотрудниками. В некоторых случаях модельными системами служили твердые пленки, которые получались путем выпаривания эфирного раствора бактериохлорофилла в вакууме, т. е. в условиях, заведомо содействующих агрегации молекул пигмента.
Хорошее совпадение спектров поглощения и спектров флуоресценции твердых пленок этого пигмента с соответствующими спектрами пурпурных бактерий указывает на наличие в живых клетках агрегированных форм бактериохлорофилла. В настоящее время считается, что пурпурные бактерии содержат три различно агрегированные формы зеленого пигмента. В соответствии с положением основных максимумов поглощения в инфракрасной части спектра при 800, 850 и 890 ∙ 10 -9 м эти формы бактериохлорофилла принято обозначать как Б800, Б850 и Б890.
Проблема множественности форм хлорофилла приобрела исключительно важное значение после того, как было установлено, что первичный процесс фотосинтеза включает в себя не одну фотохимическую реакцию, а две и что различные формы пигмента могут осуществлять самостоятельно специфические функции.

Спектры всех каротиноидов в видимой области характеризуются двумя или тремя интенсивными полосами поглощения в сине-фиолетовой части. Спектры поглощения фикоэритрина и фикоциана близки между собой, их максимумы приходятся на зеленую и желтую части спектра, располагаясь между двумя основными полосами поглощения зеленых пигментов. Это позволяет считать, что фикобилипротеиды наряду с каротиноидами являются дополнительными пигментами к хлорофиллу.
Адаптация к условиям освещения
Фикоциан и фикоэритрин как дополнительные пигменты играют существенную роль в адаптации (приспособлении) растений к условиям освещения, обеспечивая наилучшее использование солнечной энергии для фотосинтеза.
Адаптацию к качеству и интенсивности света можно наглядно видеть на примере вертикального распределения морских водорослей в зависимости от их окраски: в самых поверхностных слоях воды преобладают зеленые водоросли, в самых глубоких - красные, бурые водоросли занимают промежуточное положение. Такое распределение водорослей вызывается следующими причинами. Спектральный состав и интенсивность солнечного света, проходящего через толщу воды, претерпевают большие изменения. Поглощаясь избирательно, солнечный свет обедняется в первую очередь красными лучами, и на глубине в несколько метров он становится синевато-зеленоватым, практически лишенным красных лучей, которые полностью гасятся на глубине примерно 20 м. В связи с этим появление в процессе эволюции дополнительных к хлорофиллу пигментов, интенсивно поглощающих зеленые и желтые лучи, обеспечило красным и бурым водорослям возможность использования для фотосинтеза тех участков солнечного света, которые достигают больших глубин. Можно сказать, что наличие у бурых и красных водорослей дополнительных пигментов следует рассматривать как специальное приспособление к условиям освещения, и в первую очередь как хроматическую адаптацию к наилучшему использованию лучей преобладающей окраски, проходящих через толщу воды.
Кроме того, при прохождении света через толщу воды резко снижается и его интенсивность. Так, свет, проникающий на 20-метровую глубину, ослаблен по сравнению с освещенностью водной поверхности примерно в 20 раз. Поэтому у глубоководных морских водорослей, помимо хроматической адаптации, хорошо выражено также и приспособление к использованию света очень низкой интенсивности. Глубоководные водоросли с полным основанием могут быть отнесены к типичным «тенелюбивым» растениям с высоким содержанием пигментов, в связи с чем прямой солнечный свет повреждает их.
Что касается наземных растений, то у них первостепенное значение имеет адаптация к интенсивности света, поскольку интенсивность света подвержена колебаниям в большей мере, чем его спектральный состав. Хроматическая адаптация выражена у них слабо и проявляется в изменении соотношений между разными пигментами хлоропластов, главным образом между хлорофиллом a и хлорофиллом b, а также между хлорофиллами и каротиноидами.
Адаптация к интенсивности света у наземных растений проявляется в существовании тенелюбивых и светолюбивых типов. У тенелюбивых растений, произрастающих, например, под пологом леса, хорошо выражена адаптация к условиям слабого освещения. Как известно, затененные листья отличаются гораздо более высоким содержанием зеленых пигментов в целом, что содействует лучшему поглощению света низкой интенсивности. Кроме того, листья теневыносливых растений содержат больше хлорофилла Ь, ив этом как раз нетрудно усмотреть приспособление к несколько измененному спектральному составу света, проникающего под полог леса. В тени под нависающей сверху листвой рассеянный свет относительно обогащен коротковолновыми лучами в области 450-480 ∙ 10 -9 м, которые интенсивно поглощаются хлорофиллом 6, иногда называемым в связи с этим теневым зеленым пигментом.
Биосинтез пигментов
Исключительно большие успехи достигнуты за последние 20 лет в области изучения биосинтеза пигментов. Особенно глубоко к настоящему времени изучен биосинтез хлорофилла. С помощью меченых атомов, бумажной хроматографии и других современных методов исследования удалось почти полностью расшифровать химизм всех основных этапов формирования молекулы хлорофилла, определить их последовательность, а также выяснить условия, необходимые для протекания тех или иных реакций, и ферментные системы, катализирующие отдельные реакции. Мы не имеем возможности остановиться здесь подробно на всех реакциях биосинтетической цепи хлорофилла, опишем лишь вкратце основные этапы и приведем структурные формулы важнейших промежуточных соединений.
Начальный этап биосинтеза молекулы хлорофилла a включает ряд реакций, в результате которых из несложных органических веществ - аминокислоты гликокола (NH 2 -СН 2 -СООН) и янтарной кислоты (НООС-СН 2 -СН 2 -СООН) - возникает δ-аминолевулиновая кислота, а при конденсации двух молекул этой кислоты образуется порфобилиноген.

Один из следующих важных этапов складывается из реакций, приводящих сначала к синтезу основного структурного ядра хлорофилла путем соединения четырех молекул порфобилиногена, а затем к образованию протохлорофиллида а, имеющего уже в своем составе атом магния. После этого происходит восстановление протохлорофиллида, присоединяющего два атома водорода за счет двойной связи четвертого пиррольного кольца, и превращение его в хлорофиллид а. Для подавляющего большинства видов растений эта реакция является фотохимической и происходит только в присутствии света. Исключение составляют многие водоросли, а также некоторые представители мхов и папоротников, у которых реакция восстановления может осуществляться и в темноте.
Вслед за фотохимическим восстановлением имеет место присоединение к хлорофиллиду а фитола, которое не нуждается в присутствии света, но осуществляется при участии фермента хлорофиллазы. Присоединение фитола - это заключительный этап биосинтетической цепи, на котором хлорофиллид а превращается в хлорофилл а.
Раньше считалось, что к протохлорофиллиду сначала присоединяется не водород, а фитол, а затем уже образующийся при этом протохлорофилл восстанавливается фотохимически до хлорофилла. Получалось, что непосредственным предшественником хлорофилла следовало признать протохлорофилл. Сравнительно недавно было установлено, что хотя образование протохлорофилла в растении таким путем и не исключается, однако последний этап биосинтеза хлорофилла осуществляется главным образом через хлорофиллид, который служит основным предшественником хлорофилла.
С помощью меченых атомов показано, что хлорофилл a является в свою очередь предшественником хлорофилла b. При этом одновременно было доказано, что биосинтез хлорофилла b из хлорофилла а не нуждается в обязательном участии света и может происходить даже в полной темноте.
Хлорофилл с может образовываться в растениях, согласно предположению С. Граника, из протохлорофиллида а, поскольку оба эти соединения имеют невосстановленное четвертое пиррольное кольцо и лишены фитольного остатка.
Биосинтез каротиноидов изучен пока недостаточно полно и не все его звенья установлены окончательно. До сих пор не расшифрован химизм отдельных реакций и не выявлена природа соединений, которые служат непосредственными предшественниками различных каротиноидов. Лучше всего изучены начальные этапы, и можно считать окончательно доказанным, что биосинтез каротиноидов и фитола идет одним и тем же путем по крайней мере до образования мевалоновой кислоты.
Установлено также, что свет стимулирует биосинтез как каротиноидов, так и фитола. Хотя образование этих веществ протекает интенсивнее на свету, свет все-таки не обязателен для их биосинтеза. Дальнейшие этапы перехода от мевалоновой кислоты, имеющей в своем составе шесть углеродных атомов, к фитолу, содержащему двадцать углеродных атомов, и тем более к каротиноидам, содержащим сорок углеродных атомов, окончательно еще не установлены и трактуются по-разному.
В ходе исследования биосинтеза зеленых пигментов возникло представление о возможной связи между образованием и разрушением молекул хлорофилла. В дальнейшем с помощью изотопного метода Ф. В. Турчину, В. М. Кутюрину, А. А. Шлыку и другим удалось обнаружить и изучить процесс обновления хлорофилла. По мнению А. А. Шлыка, полученные экспериментальные данные позволяют говорить пока что о главном способе обновления хлорофилла - через распад молекул, ранее образовавшихся, и полный синтез новых. Скорость обновления может изменяться в зависимости как от вида растения, так и от условий произрастания. По предварительным данным, в течение суток обычно обновляется около 10% общего числа всех имеющихся молекул. Однако в некоторых случаях скорость обновления может быть очень низкой, граничащей с полной приостановкой этого процесса.
С помощью изотопного метода выяснена также особая роль только что синтезированных («молодых») молекул в обмене хлорофилла. Особая роль «молодых» молекул обусловлена тем, что они находятся в более реакционноспособном, более лабильном (подвижном) состоянии, чем «старые» молекулы. Причем повышенной лабильностью обладают «молодые» молекулы хлорофилла a и хлорофилла b. Об этом свидетельствуют различия в свойствах «молодых» и «старых» молекул. Например, «молодые» молекулы хлорофилла a легче экстрагируются органическими растворителями, легче теряют окраску и разрушаются при действии кислот, ультразвука или при отсутствии света, а также легче превращаются в хлорофилл b.
Биосинтез пигментов представляет собой цепь процессов, которые протекают не изолированно и самостоятельно, а находятся во взаимосвязи со всем обменом веществ и жизнедеятельностью растительного организма. Большое влияние на биосинтез пигментов оказывает также действие различных факторов внешней среды, в том числе и действие света, о чем говорилось выше. Так, образование хлорофилла возможно только при наличии кислорода в окружающей среде. Отсутствие кислорода тормозит этот процесс, как и понижение температуры. По данным Т. Н. Годнева и его сотрудников, синтез хлорофилла у некоторых вечнозеленых растений может наблюдаться при температуре не ниже -2°, а каротиноидов - не ниже -5°, в то время как способность осуществлять фотосинтез сохраняется у этих видов растений даже при - Небольшое влияние на образование хлорофилла оказывают условия минерального питания. Широко известно явление хлороза у растений (от греч. хлорос - зеленовато-желтый, бледный), когда листья бедны хлорофиллом и имеют желтоватую окраску. Наиболее часто хлороз вызывается отсутствием или недостатком в питательной среде железа, но эта связь наблюдается не всегда. Хлоротичные растения могут развиваться и при недостатке или отсутствии в окружающей среде магния, меди, серы, калия и некоторых других элементов минерального питания. Недостаток калия вызывает хлороз, так же как и избыточные количества этого элемента при недостаточно хорошем азотном питании. При недостатке азота наблюдается резкое ослабление синтеза хлорофилла. Отсутствие или недостаток магния в почве также неизбежно вызывает хлороз растений, поскольку этот элемент входит в состав молекулы хлорофилла.
Значение хлорофилла и каротина для медицины
Хлорофилл и другие пигменты независимо от их роли и значения для фотосинтеза сами по себе являются ценными органическими веществами. Очень важна близость химического состава каротина и витамина А. Каротин можно рассматривать как провитамин А, поскольку при расщеплении его молекулы пополам с присоединением кислорода образуется две молекулы этого витамина. У животных такое расщепление каротина до витамина А происходит при участии особого фермента - каротиназы, который содержится в их печени.
Все более широкое применение в медицине в последнее время находит и хлорофилл. Ценные терапевтические свойства хлорофилла, вероятно, можно объяснить тем, что по своему строению он весьма близок к красящему веществу гемоглобина крови. Поэтому нет ничего неожиданного в том, что в организме человека и животных хлорофилл содействует усиленному образованию гема гемоглобина крови, а также способствует заживлению ран. Кроме того, оказалось, что свойства хлорофилла и витаминов во многом сходны.
Хлорофилл – это термин, используемый для обозначения нескольких тесно связанных между собой зеленых пигментов, содержащихся в цианобактериях и хлоропластах водорослей и растений. Название происходит от греческих слов χλωρός, хлорос («зеленый») и φύλλον, phyllon («лист»). Хлорофилл является чрезвычайно важной биомолекулой, имеет решающее значение в процессе фотосинтеза, который позволяет растениям поглощать энергию света. Хлорофилл наиболее интенсивно поглощает свет в синей части спектра электромагнитного излучения, а также в красной части. С другой стороны, хлорофилл плохо поглощает зеленую и близкую к зеленой части спектра, которые он отражает, поэтому хлорофилл-содержащие ткани имеют зеленый цвет. Хлорофилл впервые был выделен и назван Жозефом Бьенеме Каванту и Пьером Жозефом Пеллетье в 1817 году.
Хлорофилл и фотосинтез
Хлорофилл имеет жизненно важное значение для фотосинтеза, который позволяет растениям поглощать энергию света. Молекулы хлорофилла специально расположены внутри и вокруг фотосистем, которые встроены в мембраны тилакоидов хлоропластов. В этих комплексах, хлорофилл выполняет две основные функции. Функция подавляющего большинства хлорофилла (до нескольких сотен молекул в фотосистеме) состоит в том, чтобы поглощать свет и передавать энергию света путем резонансного переноса энергии к конкретной паре хлорофилла в реакционном центре фотосистем. Две принятые в настоящее время единицы фотосистем – фотосистема II и фотосистема I, которые имеют свои собственные различные реакционные центры, названные Р680 и Р700, соответственно. Эти центры названы по длине волны (в нанометрах) их максимального поглощения в красном спектре. Идентичность, функциональность и спектральные свойства хлорофилла в каждой фотосистеме различны и определяются друг другом и белковой структурой, окружающей их. После извлечения из белка в растворителе (таком, как ацетон или метанол), пигменты хлорофилла могут быть разделены на хлорофилл а и б. Функция реакционного центра хлорофилла – поглощать энергию света и переносить её на другие части фотосистемы. Поглощенная энергия фотона передается электрону в процессе, называемом разделение зарядов. Удаление электрона из хлорофилла является реакцией окисления. Хлорофилл жертвует электроном с высокой энергией ряду молекулярных промежуточных продуктов, называемых цепью переноса электронов. Заряженный реакционный центр хлорофилла (P680 +) затем восстанавливается обратно в основное состояние, принимая электрон, отделенный от воды. Электрон, который восстанавливает Р680 +, в конечном счете, происходит от окисления воды в О2 и Н + через несколько промежуточных продуктов. В ходе этой реакции, фотосинтезирующие организмы, такие как растения, производят O2 газ, который является источником практически всего O2 в атмосфере Земли. Фотосистема I обычно работает последовательно с фотосистемой II; таким образом, P700 + фотосистемы I обычно восстанавливается, когда он принимает электрон, через множество промежуточных в тилакоидной мембране, при помощи электронов, которые приходят, в конечном счете, от фотосистемы II. Реакции переноса электронов в мембранах тилакоидов сложны, и источник электронов, используемый для восстановления P700 +, может меняться. Электронный поток, который создается пигментами реакционного центра хлорофилла, используется для накачки ионов Н + через мембрану тилакоида, настраивая хемиосмотической потенциал, используемый, главным образом, в производстве АТФ (накопленная химическая энергия), или в восстановлении NADP + в NADPH. НАДФ является универсальным агентом, используемым для восстановления СО2 в сахара, а также в других биосинтетических реакциях. РЦ хлорофилл-белковые комплексы способны непосредственно поглощать свет и разделять заряды без помощи других хлорофилловых пигментов, но вероятность этого при заданной интенсивности света мала. Таким образом, другие хлорофиллы фотосистемы и антенные пигментные белки кооперативно поглощают и переносят световую энергию к реакционному центру. Кроме хлорофилла а, существуют и другие пигменты, называемые вспомогательными пигментами, которые имеют место в этих антенных пигмент-белковых комплексах.
Химическая структура
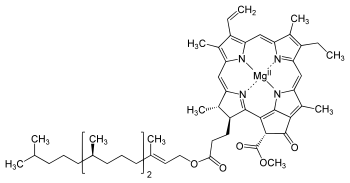 Хлорофилл представляет собой хлориновый пигмент, который структурно подобен и производится в рамках того же пути метаболизма, что и другие порфириновые пигменты, такие как гем. В центре кольца хлорина находится ион магния. Это было обнаружено в 1906 году, и впервые магний был обнаружен в живой ткани. Хлориновое кольцо может иметь несколько различных боковых цепей, как правило, включающих длинную цепь фитола. Есть несколько различных форм, которые встречаются в природе, но наиболее широко у наземных растений распространена форма хлорофилл а. После первоначальной работы, проделанной немецким химиком Ричардом Вильштеттером с 1905 по 1915 годы, Ганс Фишер определил общую структуру хлорофилла а в 1940 г. К 1960 г., когда большая часть стереохимии хлорофилла а была известна, Вудворд опубликовал полный синтез молекулы. В 1967 году, последнее оставшееся стереохимическое объяснение было дано Яном Флемингом, а в 1990 году Вудворд и соавторы опубликовали обновленный синтез. Было объявлено, что хлорофилл е присутствует в цианобактериях и других оксигенных микроорганизмах, которые образуют строматолиты, в 2010 году. Молекулярная формула C55H70O6N4Mg и структура (2-формил)-хлорофил были выведены на основе ЯМР, оптического и масс-спектров.
Хлорофилл представляет собой хлориновый пигмент, который структурно подобен и производится в рамках того же пути метаболизма, что и другие порфириновые пигменты, такие как гем. В центре кольца хлорина находится ион магния. Это было обнаружено в 1906 году, и впервые магний был обнаружен в живой ткани. Хлориновое кольцо может иметь несколько различных боковых цепей, как правило, включающих длинную цепь фитола. Есть несколько различных форм, которые встречаются в природе, но наиболее широко у наземных растений распространена форма хлорофилл а. После первоначальной работы, проделанной немецким химиком Ричардом Вильштеттером с 1905 по 1915 годы, Ганс Фишер определил общую структуру хлорофилла а в 1940 г. К 1960 г., когда большая часть стереохимии хлорофилла а была известна, Вудворд опубликовал полный синтез молекулы. В 1967 году, последнее оставшееся стереохимическое объяснение было дано Яном Флемингом, а в 1990 году Вудворд и соавторы опубликовали обновленный синтез. Было объявлено, что хлорофилл е присутствует в цианобактериях и других оксигенных микроорганизмах, которые образуют строматолиты, в 2010 году. Молекулярная формула C55H70O6N4Mg и структура (2-формил)-хлорофил были выведены на основе ЯМР, оптического и масс-спектров.
Измерение содержания хлорофилла
Измерение поглощения света осложняется растворителем, используемым для извлечения хлорофилла из растительного материала, который влияет на полученные значения. В диэтиловом эфире, хлорофилл а имеет приблизительные максимумы абсорбции 430 нм и 662 нм, в то время как хлорофилл б имеет приблизительные максимумы 453 нм и 642 нм. Пики поглощения хлорофилла а составляют 665 нм и 465 нм. Хлорофилл флуоресцирует при 673 нм (максимум) и 726 нм. Пиковый молярный коэффициент поглощения хлорофилла а превышает 105 М-1 см-1, и является одним из самых высоких для малых молекул органических соединений. В 90% ацетона-воды, пиковые длины волн поглощения хлорофилла а составляют 430 нм и 664 нм; пики для хлорофилла б – 460 нм и 647 нм; пики для хлорофилла c1 – 442 нм и 630 нм; пики для хлорофилла с2 – 444 нм и 630 нм; пики для хлорофилла d – 401 нм, 455 нм и 696 нм. Измерив поглощение света в красном и дальнем красном спектрах, можно оценить концентрацию хлорофилла в листе. Коэффициент эмиссии флуоресценции может быть использован для измерения содержания хлорофилла. Возбуждая флуоресценцию хлорофилла «а» при более низкой длине волны, отношение флуоресцентной эмиссии хлорофиллов при 705 нм +/- 10 нм и 735 нм +/- 10 нм может обеспечить линейную зависимость содержания хлорофилла по сравнению с химическими испытаниями. Отношение F735 / F700 обеспечило значение корреляции r2 0,96 по сравнению с химическими испытаниями в диапазоне от 41 мг м-2 до 675 мг м-2. Гительзон также разработал формулу для прямого считывания содержания хлорофилла в мг м-2. Формула обеспечила надежный метод измерения содержания хлорофилла от 41 мг м-2 до 675 мг м-2 со значением корреляции r2 0,95.
Биосинтез
В растениях, хлорофилл может быть синтезирован из сукцинил-СоА и глицина, хотя непосредственным предшественником хлорофилла а и б является протохлорофиллид. У покрытосеменных растений, последний шаг, превращение протохлорофиллида в хлорофилл, зависит от освещенности, и такие растения являются бледными, если выращены в темноте. Несосудистые растения и зеленые водоросли имеют дополнительный фермент, независимый от света, и способны зеленеть в темноте. Хлорофилл связывается с белками и может передавать поглощенную энергию в нужном направлении. Протохлорофиллид встречается, главным образом, в свободной форме, и в условиях освещенности действует в качестве фотосенсибилизатора, образуя высокотоксичные свободные радикалы. Следовательно, растениям необходим эффективный механизм регулирования количества предшественника хлорофилла. У покрытосеменных растений, это делается на стадии аминолевулиновой кислоты (ALA), одного из промежуточных соединений в пути биосинтеза. Растения, которые питаются ALA, накапливают высокие и токсические уровни протохлорофиллида; так же делают мутанты с поврежденной системой регулирования.
Хлороз
Хлороз – это состояние, при котором листья производят недостаточное количество хлорофилла, что делает их желтыми. Хлороз может быть вызван питательным дефицитом железа, называемого хлорозом железа, либо нехваткой магния или азота. РН почвы иногда играет роль в хлорозе, вызванном питанием; многие растения приспособлены расти в почвах с определенными уровнями рН и их способности поглощать питательные вещества из почвы могут зависеть от этого. Хлороз также может быть вызван патогенными микроорганизмами, включая вирусы, бактерии и грибковые инфекции, или сосущих насекомых.
Дополнительное поглощение света антоцианов с хлорофиллом
Антоцианы – это другие растительные пигменты. Паттерн абсорбции, отвечающий за красный цвет антоцианов, может дополнять зеленый хлорофилл в фотосинтетически активных тканях, таких как молодые листья Quercus coccifera. Он может защищать листья от нападений со стороны травоядных, которые могут быть привлечены зеленым цветом.
Использование хлорофилла
Кулинарное использование
 Хлорофилл зарегистрирован в качестве пищевой добавки (краситель), и его номер E140. Повара используют хлорофилл для окрашивания различных пищевых продуктов и напитков в зеленый цвет, такие как макароны и абсент. Хлорофилл не растворяется в воде, и его сначала смешивают с небольшим количеством растительного масла, чтобы получить желаемый раствор.
Хлорофилл зарегистрирован в качестве пищевой добавки (краситель), и его номер E140. Повара используют хлорофилл для окрашивания различных пищевых продуктов и напитков в зеленый цвет, такие как макароны и абсент. Хлорофилл не растворяется в воде, и его сначала смешивают с небольшим количеством растительного масла, чтобы получить желаемый раствор.
Польза для здоровья
Хлорофилл способствует укреплению органов кроветворения, обеспечивая профилактику анемии и обилие кислорода в организме. Его антиоксидантная активность оказывает благотворное влияние на различные медицинские состояния, такие как рак, бессонница, стоматологические заболевания, синусит, панкреатит и камни в почках. Хлорофилл способствует нормальной свертываемости крови, заживлению ран, гормональному балансу, дезодорации и детоксикации организма и способствует здоровью пищеварительной системы. Он благотворно влияет на окисление и воспалительные заболевания, такие как артрит и фибромиалгия. Он проявляет омолаживающие и противомикробные свойства и способствует укреплению иммунной системы организма.
Общее
Хлорофилл является пищевым продуктом, содержащим большое количество питательных веществ. Он является хорошим источником витаминов, таких как витамин А, витамин С, витамин Е, витамин К и бета-каротин. Он богат антиоксидантами, жизненно важными минералами, такими как магний, железо, калий, кальций и незаменимые жирные кислоты.
Эритроциты
Хлорофилл помогает в восстановлении и пополнении запасов красных кровяных клеток. Он работает на молекулярном и клеточном уровнях и обладает способностью к регенерации нашего тела. Он богат живыми ферментами, которые способствуют очищению крови и повышают способность крови переносить больше кислорода. Он является строителем крови и также эффективен против анемии, которая вызвана дефицитом красных кровяных клеток в организме.
Рак
Хлорофилл эффективен против рака, например, рака толстой кишки человека, и стимулирует индукцию апоптоза. Он обеспечивает защиту от широкого спектра канцерогенов, присутствующих в воздухе, приготовленном мясе и зерне. Исследования показали, что хлорофилл помогает в сдерживании желудочно-кишечной абсорбции вредных токсинов, также известных как афлатоксины, в организме. Хлорофилл и его производное хлорофиллин ингибируют метаболизм этих прокарциногенов, которые могут нанести ущерб ДНК, а также привести к раку печени и гепатиту. Дальнейшие исследования, проведенные в этой связи, демонстрируют химио-профилактическое действие хлорофилла, приписывая ему антимутагенные свойства. Другое исследование показало эффективность диетического хлорофилла в качестве фитохимического соединения, сокращающего онкогенез.
Антиоксидант
Хлорофилл обладает сильной антиоксидантной активностью, наряду со значительным количеством необходимых витаминов. Эти эффективные поглотители радикалов помогают нейтрализовать вредные молекулы и защитить от развития различных заболеваний и повреждений в результате окислительного стресса, вызванного свободными радикалами.
Артрит
Противовоспалительные свойства хлорофилла являются полезными для лечения артрита. Исследования показали, что хлорофилл и его производные мешают росту воспаления, вызванного воздействием бактерий. Этот защитный характер хлорофилла делает его мощным ингредиентом для приготовления фитосанитарных продуктов для лечения болезненных медицинских состояний, таких как фибромиалгия и артрит.
Детоксикация
Хлорофилл имеет очищающие свойства, которые помогают в детоксикации организма. Обилие кислорода и здорового потока крови из-за хлорофилла в организме способствует избавлению от вредных примесей и токсинов. Хлорофилл образует комплексы с мутагенами и обладает способностью связывать и вымывать токсичные химические вещества и тяжелые металлы, такие как ртуть, из организма. Он способствует детоксикации и возрождению печени. Он также эффективен в снижении вредного воздействия излучений и помогает устранить пестициды и отложения препаратов из организма.
Анти-старение
Хлорофилл помогает в борьбе с последствиями старения и поддерживает здоровье тканей, из-за богатства антиоксидантов и присутствия магния. Он стимулирует ферменты, действующие против старения, и способствует здоровью и молодости кожи. В дополнение к этому, витамин K, присутствующий в нем, очищает и омолаживает надпочечники и улучшает функции надпочечников в организме.
Пищеварительная система
Хлорофилл способствует здоровому пищеварению путем поддержания кишечной флоры и стимулирования перистальтики кишечника. Он действует как естественный препарат для желудочно-кишечного тракта и помогает в восстановлении поврежденных тканей кишечника. Диеты с дефицитом зеленых овощей и включающие, в основном, красное мясо, представляют повышенную опасность нарушений толстой кишки. Согласно исследованиям, хлорофилл облегчает очищение толстой кишки путем ингибирования цитотоксичности, вызванной диетическим гемом и предотвращает распространение колоноцитов. Он эффективен при запорах и уменьшает дискомфорт, вызванный газами.
Бессонница
Хлорофилл оказывает успокаивающее действие на нервы и помогает в снижении симптомов бессонницы, раздражительности и нервной общей усталости организма.
Антимикробные свойства
Хлорофилл обладает эффективными противомикробными свойствами. Недавние исследования показали, что лечебный эффект щелочного раствора на основе хлорофилла в борьбе с заболеванием, называемым Candida Albicans, инфекцией, вызванной разрастанием дрожжей кандида, уже присутствует в небольших количествах в организме человека.
Иммунитет
Хлорофилл способствует укреплению клеточных стенок и общей иммунной системы организма из-за его щелочного характера. Анаэробные бактерии, которые способствуют развитию заболеваний, не могут выжить в щелочной среде хлорофилла. Наряду с этим, хлорофилл является оксигенатором, который поощряет способность организма бороться с заболеваниями и повышает уровень энергии и ускоряет процесс заживления.
Дезодорирующие свойства
Хлорофилл проявляет дезодорирующие свойства. Он является эффективным средством для борьбы с неприятным запахом изо рта и используется в составе жидкостей для полоскания рта. Плохое здоровье пищеварительной является одной из основных причин неприятного запаха изо рта. Хлорофилл выполняет двойное действие, устраняя запах изо рта и горла, а также стимулируя здоровье пищеварительной системы путем очищения толстой кишки и потока крови. Дезодорирующий эффект хлорофилла также эффективно воздействует на раны, имеющие неприятный запах. Его вводят перорально пациентам, страдающим от колостомы и нарушений обмена веществ, таких как триметиламинурия, для снижения фекального и мочевого запаха.
Заживление ран
Исследования показывают, что местное применение растворов хлорофилла является эффективным при лечении ран и ожогов. Он помогает снижать местные воспаления, укрепляет ткани организма, помогает убивать микробов и повышает устойчивость клеток против инфекций. Он предотвращает рост бактерий, дезинфицируя окружающую среду, делая её враждебной для роста бактерий, и ускоряет заживление. Хлорофилл также весьма эффективен при лечении хронических варикозных язв.
Кислотно-щелочное соотношение
Потребление продуктов, богатых хлорофиллом, помогает сбалансировать кислотно-щелочной баланс в организме. Магний, присутствующий в нем, является мощной щелочью. Поддерживая соответствующую щелочность и уровни кислорода в организме, хлорофилл предотвращает развитие среды для роста патогенных микроорганизмов. Магний, присутствующий в хлорофилле, также играет важную роль в поддержании здоровья сердечно-сосудистой системы, функционировании почек, мышц, печени и головного мозга.
Сильные кости и мышцы
Хлорофилл способствует формированию и поддержанию крепких костей. Центральный атом молекулы хлорофилла, т.е. магний, играет важную роль в здоровье костей, наряду с другими необходимыми питательными веществами, такими как кальций и витамин D. Он также вносит вклад в тонус, сокращение и расслабление мышц.
Свертывание крови
Хлорофилл содержит витамин К, который имеет жизненно важное значение для нормального свертывания крови. Он используется в натуропатии для лечения носовых кровотечений и для женщин, страдающих от анемии и тяжелого менструального кровотечения.
Камни в почках
Хлорофилл способствует предотвращению образования камней в почках. Витамин К присутствует в виде эфирных соединений хлорофилла в моче и помогает в снижении роста кристаллов оксалата кальция.
Синусит
Хлорофилл эффективен в лечении различных респираторных инфекций и других заболеваний, таких как простуда, ринит и синусит.
Гормональный баланс
Хлорофилл полезен в поддержании полового гормонального баланса у мужчин и женщин. Витамин Е, присутствующий в хлорофилле, помогает стимулировать выработку тестостерона у мужчин и эстрогена у женщин.
Панкреатит
Хлорофилл вводят внутривенно при лечении хронического панкреатита. Согласно результатам исследования, проведенного в связи с этим, он помогает в снижении лихорадки и уменьшает боль в животе и дискомфорт, вызванный панкреатитом, не вызывая никаких побочных эффектов.
Гигиена полости рта
Хлорофилл помогает в лечении стоматологических проблем, таких как пиорея. Он используется для лечения симптомов инфекции полости рта и успокаивает воспаленные и кровоточащие десна.
Источники хлорофилла
Не очень трудно включить хлорофилл в ежедневный рацион, так как почти все зеленые растения богаты хлорофиллом а, и многие овощи, которые являются неотъемлемой частью нашей пищи, содержат хлорофилл а, а также хлорофилл б. Потребление овощей, таких как руккола, ростки пшеницы, лук-порей, зеленые бобы и темно-зеленые листовые овощи, такие как петрушка, капуста, кресс-салат, листовая свекла и шпинат, обеспечивают естественный хлорофилл для организма. Другие источники включают капусту, синие зеленые водоросли, такие как хлорелла и спирулина. Приготовление пищи разрушает содержащиеся в ней хлорофилл и магний, поэтому сырые или приготовленные на пару овощи являются более полезными.
Предостережения
Несмотря на клиническое использование в течение многих лет, токсические эффекты естественного хлорофилла в обычных дозах не были известны. Тем не менее, хлорофилл может вызвать некоторое изменение цвета языка, мочи или кала при пероральном введении. Наряду с этим, хлорофилл также может вызвать легкое жжение или зуд при местном применении. В редких случаях, передозировка хлорофиллом может привести к диарее, спазмам в животе и поносу. При таких симптомах, целесообразно обратиться за медицинской помощью. Беременные или кормящие женщины должны воздерживаться от использования коммерчески доступного хлорофилла или добавок хлорофилина из-за отсутствия доказательств безопасности.
Лекарственные взаимодействия
Пациентам, проходящим гваяковую пробу на скрытую кровь, следует избегать перорального использования хлорофиллина, так как это может привести к ложноположительному результату.
Резюме
Хлорофилл обеспечивает энергию солнца в концентрированной форме для нашего тела и является одним из самых полезных питательных веществ. Он повышает уровень энергии и усиливает общее состояние благополучия. Он также полезен при ожирении, сахарном диабете, гастрите, геморрое, астме и кожных заболеваниях, таких как экзема. Он помогает в лечении высыпаний и в борьбе с инфекциями кожи. Потребление хлорофилла профилактически также предотвращает неблагоприятные последствия хирургического вмешательства и его рекомендуется вводить до и после операции. Содержание в нем магния помогает в поддержании кровотока в организме и поддерживает нормальный уровень кровяного давления. Хлорофилл в целом улучшает клеточный рост и восстанавливает здоровье и бодрость в организме.
:Tags
Список использованной литературы:
Meskauskiene R; Nater M; Goslings D; Kessler F; op den Camp R; Apel K. (23 October 2001). «FLU: A negative regulator of chlorophyll biosynthesis in Arabidopsis thaliana». Proceedings of the National Academy of Sciences. 98 (22): 12826–12831. Bibcode:2001PNAS…9812826M. doi:10.1073/pnas.221252798. JSTOR 3056990. PMC 60138free to read. PMID 11606728
Adams, Jad (2004). Hideous absinthe: a history of the devil in a bottle. United Kingdom: I.B.Tauris, 2004. p. 22. ISBN 1860649203.

Основным пигментом зеленых растений является молекула хлорофилла, участвующая в процессе поглощения света. Высшие растения содержат две формы хлорофилла: хлорофилл а и хлорофилл b. Структура хлорофилла а (рис. 40) установлена Вилыптеттером и Фишером и подтверждена в 1960 г. Вудвордом, осуществившим полный синтез хлорофилла а.
В основе молекулы хлорофилла лежит плоское порфириновое кольцо, в центре которого находится ион атома магния, координационно соединенный с атомами азота порфиринового кольца.
Плоская структура порфиринового кольца обусловлена сопряженными двойными и простыми связями -электронов между атомами углерода и азота. Эти электроны «делокализованы», т. е. равномерно распределены вдоль «периферии» порфиринового кольца (выделенная точками область на рис. 40). Изменение состояния движения -электронов в кольце требует сравнительно небольшой энергии. Поэтому спектр поглощения света молекулой хлорофилла лежит в красной области. Дипольный электрический момент перехода в возбужденное состояние находится в плоскости порфиринового кольца.
Кроме порфиринового кольца молекула хлорофилла имеет длинную гидрофобную цепь - «хвост», в состав которого входят 20 углеродных атомов. Эта боковая цепь представляет собой остаток спирта фитола. Хлорофилл b отличается от хлорофилла а тем, что в последнем группа - заменена группой - СНО. Таким образом, хлорофилл b содержит на один атом кислорода больше и на два атома водорода меньше, чем хлорофилл а.
Спектры поглощения обеих форм хлорофилла приведены на рис. 41. Максимумы полос поглощения хлорофилла а лежат в областях длин волн К да 700 нм (красная) и К да 440 нм (фиолетовая), максимумы полос поглощения хлорофилла b - в областях длин волн 660 и 460 нм.
Максимальная интенсивность солнечного света, достигающего земной поверхности, приходится на сине-зеленую и зеленую области длин волн (450-550 нм). Оказывается, что именно в этих областях поглощение света молекулами хлорофилла минимально.
Хлорофилл а найден у всех зеленых растений и водорослей. Хлорофилл b отсутствует у многих водорослей. Эти водоросли иногда содержат другие разновидности хлорофилла: с и d. Фотосинтезирующие бактерии, не выделяющие кислород, хлорофилла а не содержат. В их состав обычно входит особый тип хлорофилла- бактериохлорофилл.
Как указывалось выше, кроме молекул хлорофилла многие фотосинтезирующие клетки содержат также пигментные молекулы,

Рис. 40. Структурные формулы хлорофилла а и хлорофилла b.
поглощающие свет в других областях спектра и придающие организмам различную окраску. Эти молекулы расширяют спектральную область света, используемую при фотосинтезе. Кроме того, каротиноиды предохраняют хлорофилл от необратимого фотоокисления кислородом.
Структурные формулы одного из каротинов и фикоцианобилина указаны на рис. 42. Каротины имеют длинные полиизопреновые цепи сопряженных двойных и одиночных связей. На каждом конце молекулы находятся циклогексановые кольца. Фикоциашшы, входящие в состав синезеленых водорослей, содержат четыре пи-рольных кольца. Они могут образовывать комплексы со специфическими белками.
На рис. 43 представлена схема первых энергетических уровней молекулы хлорофилла а. В основном состоянии молекула имеет нулевой спин. Все возбужденные состояния с нулевым спином называются синглетными (S). У молекулы возможны также возбужденные состояния со спином единица (в единицах h). Они называются триплетными (Т). Время жизни первого синглетного состояния . Время жизни нулевого триплетного состояния .

Рис. 41. Спектры поглощения света хлорофиллом а (1) и хлорофиллом b (2).
Под действием света в молекуле осуществляются только переходы в синглетные возбужденные состояния. Если молекулы хлорофилла при поглощении света переходят в возбужденные состояния с энергиями, превышающими энергию первого возбужденного состояния то вследствие безыэлучательных процессов за время 10-12 - 10-13 с они переходят в первое синглетное нозбужденное состояние отдавая избыток энергии растворителю.
Из синглегного состояния за время происходит переход в основное состояние с излучением света ( нм). Это явление называется флуоресценцией. Имеется также небольшая вероятность безызлучательного перехода молекулы из нозбужденного состояния в триплетное возбужденное состояние Из-за слабого взаимодействия спина с электромагнитной волной время жизни триплетного состояния по отношению к излучению света X я» 930 нм при переходе в основное синглетное состояние сравнительно велико . Большая длительность премени жизни триплетного состояния обусловлена маловероятным процессом изменения спина молекулы от единицы до нуля.
Растворы, содержащие молекулы пигментов только одного типа (хлорофилл b, хлорофилл а, каротиноиды и др.), при низких температурах имеют характерные спектры флуоресценции, соответствующие квантовым переходам -электронов из нижайших синлетных возбужденных состояний в основное синглетное состояние молекулы. Наряду с основным излучением наблюдается слабое, медленно спадающее и более длинноволновое излучение, соответствующее переходам с нижайших триплетных состояний этих молекул в основное синглетное состояние.
В связи с тем что электронные переходы в пигментных молекулах сопровождаются изменением многих низкочастотных колебательных состояний молекул и окружающей среды, их полосы поглощения и люминесценции имеют значительную ширину.
При исследовании флуоресценции пигментов, входящих в состав

Рис. 42. Структурные формулы фотосинтезирующих пигментов: а - бета-каратин; б - фикоцианобилин.
хлоропластов, наблюдается только флуоресценция хлорофилла а. Более коротковолновая флуоресценция хлорофилла 6 и других пигментных молекул не обнаруживается даже в том случае, когда хлоропласт освещается светом с длиной волны, совпадающей с длиной волны спектра поглощения соответствующего пигмента.
Таким образом, основная масса пигментных молекул выполняет роль светособирающих систем (антенн). Пигментные молекулы в хлоропластах образуют ансамбли упорядоченно расположенных молекул .
Отмеченные выше свойства флуоресценции хлоропластов указывают, что в таких ансамблях происходит сравнительно быстрая (10-11 - 10-12 с) миграция энергии синглетного возбуждения по молекулам пигмента к молекулам хлорофилла а.
Квантовая теория систем слабо взаимодействующих одинаковых молекул показывает , что вследствие резонансного взаимодействия между возбужденной и невозбужденной молекулами в системе возникают коллективные бестоковые возбужденные состояния - экситоны, переносящие возбуждение от одних мест системы к другим. Резонансное взаимодействие с увеличением расстояния убывает сравнительно медленно (как ) и может проявляться даже на расстояниях порядка 50 А.
Когда экситон, перемещаясь по системе пигментных молекул, достигает молекулы хлорофилла а, имеющей более низкий уровень возбуждения, он переводит ее в возбужденное состояние,

Рис. 43. Схема синглетных (S t) и триплетных (71,) энергетических уровней молекулы хлорофилла а.
Прямые отрепки соответствуют поглощению, волнистые стрелки - флуоресценции; цифры указывают длины волн в нанометрах.
отдавая избыток энергии тепловому резервуару. Такая небольшая потеря энергии исключает обратный переход энергии возбуждения с молекул хлорофилла а к пигментным светособирающим молекулам.
Молекула хлорофилла а, получившая энергию от светособирающих молекул, отдает ее в виде излучения света - флуоресценции. Это явление хорошо изучено при исследовании люминесценции молекулярных кристаллов, содержащих примесные молекулы с энергией возбуждения более низкой, чем энергия возбуждения молекул основного вещества, и называется сенсибилизированной люминесценцией.
Некоторое время считали, что молекулами, воспринимающими энергию возбуждения от светособирающих молекул, являются особые молекулы хлорофилла а. Теперь установлено (см. п. 17.2), что эту роль выполняют в хлоропластах и хроматофорах особые фотосинтезирующие реакционные центры, в состав которых входит несколько молекул хлорофилла. Эти молекулы в реакционном центре образуют своеобразный комплекс, выступающий как единое целое со своим спектром возбужденных состояний. При этом энергия нижайшего из них меньше энергии отдельной молекулы хлорофилла. Установлено, что число реакционных центров в мембране значительно меньше числа светособирающих молекул (1/400).
Фотосинтезирующие реакционные центры (ловушки экситонов) входят в состав фотосинтезирующих систем (ФС), в которых осуществляются световые реакции фотосинтеза. Фотосинтезирующие системы наряду с реакционными центрами, воспринимающими энергию света, содержат ряд других молекул - ферменты, белки, липиды, липопротеиды, которые участвуют в организации фотосинтезирующей системы и в выполнении ею световой части биохимических реакций. Фотосинтезирующие системы сравнительно жестко вмонтированы в мембраны тилакоидов.
С точки зрения исследования первичных процессов фотосинтеза на молекулярном уровне особый интерес представляет изучение организации пигментных слоев и структуры фотосинтезирующих систем, в частности изучение реакционных центров, входящих в их состав.